 Kortikale
Verarbeitungsströme
Kortikale
Verarbeitungsströme
Strahlenbrechung: Das Auge enthält ein Linsensystem, das alle von einem Punkt ausgehende Strahlen auf einen Punkt der Netzhaut ("Retina") zusammenführt und so auf dieser ein umgekehrtes Bild der Umwelt projiziert. Am stärksten lichtbrechend ist die Hornhaut (Cornea) zur Veränderung der Brennweite lässt sich die dahinter liegende Linse verformen. Kurzfristige Änderungen der Lichtintensität können durch eine Veränderung der Pupillengröße durch eine Veränderung der Iris ausgeglichen werden. Bei längerfristiger Änderung der Lichtverhältnisse kommt es zu einer Anpassung der Photorezeptoren an die höchste Leuchtdichte im Gesichtsfeld (Adaptation). Das Licht unterschiedlicher Wellenlängen wird unterschiedlich stark gebrochen (chromatische Aberration), weshalb sich das Auge stets auf die Brennweite für grünes Licht einstellt und Farben räumlich zunächst nur sehr grob ausgewertet werden.
In den lichtempfindlichen Zellen der Netzhaut befinden sich in der Membran G-Protein gekoppelte Rezeptorproteine, die Rhodopsin-Moleküle, die aus aus Vitamin A (Retinal) und einem Proteinanteil (Opsin) bestehen. Fällt ein Photon auf das am C-Atom 11 geknickte Vitamin-A-Molekül (11-cis-Retinal) klappt dieses auf und wird gerade (all-trans-Retinal). Der Proteinanteil verändert seine Konformation und aktiviert in den Photorezeptoren das Transducin, ein G-Protein. Dieses löst eine signalverstärkende Enzymkaskade aus, bei Wirbeltieren (Vertebraten) hyperpolarisiert die Rezeptormembran, bei Evertebraten depolarisiert die Photorezeptormembran, das Ende der visuellen Signaltransduktionskaskade ist erreicht. Dieses elektrische Signal wird von den weiteren Zellen der Retina ausgewertet.
Das Signal wird dabei in der Retina horizontal und vertikal weitergeleitet. Horizontal regulieren Horizontalzellen und Amakrine-Zellen das Signal, vertikal wird es von Bipolarzellen an die Ganglienzellen weitergeleitet.
Die Ganglienzellen sind Neurone, deren Axone den Sehnerv bilden. Da die Retina von Wirbeltieren invers gebaut ist, die Photorezeptoren also vom Licht abgewendet sind, müssen die Axone der Ganglienzellen durch einen Punkt, den Blinden Fleck, das Auge als Sehnerv verlassen.
Schon auf der Retina wird das Signal der Photorezeptoren ausgewertet. So sorgen am Photorezeptor schon die Horizontalzellen für eine Kantenverstärkung durch die laterale Inhibition. Der zentrale Bereich der Retina ist außerdem räumlich höher aufgelöst, nur hier hat jeder Photorezeptor eine eigene Ganglienzelle. Im Durchschnitt kommen auf jede Ganglienzelle etwa 300 Photorezeptoren, in der Peripherie der Retina kann das Verhältnis bis zu 3000 Photorezeptoren pro Ganglienzelle betragen, weshalb man von dendritischen Feldern sprechen muss.
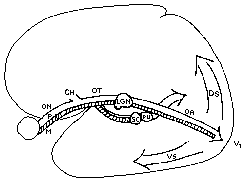
Die Fasern der Sehnerven der beider Augen werden im Chiasma opticum (Sehnervenkreuzung) so geteilt, dass die Information der linken und rechten Hemisphäre jeweils in das linke und rechte Gehirn weitergeleitet werden. Der so geteilte Sehnerv wird als Tractus opticus bezeichnet und übermittelt die Information in das Mittelhirn (Mesencephalon) in den jeweiligen Colliculus superior und das Corpus geniculatum lateralis (CGL). Vom CGL aus strahlen die Sehstrahlen (Radiatio optica) in den primären visuellen Kortex (V1) aus. Einige Zellen des CGL strahlen auch direkt in höhere Gehirnareale aus, wie das für die Bewegungserkennung zuständige visuelle Kortexareal V5 (auch mediotemporaler Kortex oder kurz MT genannt) - diese Signale dienen vermutlich zur direkten Kontrolle der Bewegungswahrnehmung. Abhängig von der Größe der Zellkörper im CGL spricht man auch vom magnozellulären (groß) und parvozellulären (klein) Verarbeitungsweg. Beide Wege haben unterschiedliche Funktionen und haben unterschiedliche Ganglienzelltypen (M- und P-Ganglienzellen, in der Literatur werden die P-Zellen midget ganglion cells, die M-Zellen parasol ganglion cells genannt). Die bei Säugetieren gefundenen W-Zellen lassen sich bei Primaten nicht nachweisen. Bisher wird angenommen, dass die großen Zellkörper vor allem für Bewegungswahrnehmung und Objektlokalisation, die kleinen vor allem für Beschaffenheit, Struktur und Farbe zuständig sind. Im CGL wurde inzwischen, zusätzlich zum magnozellulären und parvozellulären Verarbeitungsweg, ein dritter Verarbeitungsweg gefunden. Wegen der nur kleinen, vereinzelt zwischen den Schichten vorkommenden Zellen bezeichnet man ihn als den koniozellulären Verarbeitungsweg (von griechisch konios, Staub). Er dient wohl zur Verifizierung und Falsifizierung der in V1 bis V3 gewonnenen Informationen des magno- und parvozellulären Weges und ist daher direkt mit höheren Hirnarealen verschaltet (z. B. mit V5 für das Bewegungssehen).
Kortikale
VerarbeitungsströmeIm visuellen Kortexareal V1 wird vor allem eine Kantenerkennung durchgeführt. Diese Informationen werden in das Areal V2 und V3 des visuellen Kortex weitergeleitet. Ab hier teilen sich die Verarbeitungswege in einen parietalen (dorsalen, entlang des Scheitels zentral nach vorne) und einen temporalen (ventralen, zur Schläfe hin gerichteten) Verarbeitungsstrom. Diese haben unterschiedliche Funktionen. So dient der Verarbeitungsstrom zur Schläfe hin gerichtet vor allem der Objekterkennung (daher auch Was-Strom genannt), der am Scheitel entlang laufende Verarbeitungsstrom der Bewegungs- und Entfernungsbestimmung (daher auch Wo-Strom genannt). Durch diese parallele Verarbeitung wird eine enorm hohe Verarbeitungsgeschwindigkeit erreicht. Innerhalb von nur 150ms ist die gesamte Objekterkennung abgeschlossen, was zeitlich (nach der Phototransduktion) auf lediglich 5-10 neuronale Verarbeitungsschritte schließen lässt. Beide Verarbeitungsströme treffen im Stirnlappen (Frontal) erneut zusammen, womit die visuelle Wahrnehmung durch eine Objektkategorisierung und eine räumliche Bestimmung (Größe, Entfernung, Bewegung) abgeschlossen wird. Das Sehen eines Objektes erfolgt also zeitgleich mit seiner Wahrnehmung! Entlang dieser kortikalen Verarbeitungswege wird die räumliche Anordnung der retinalen Ganglienzellen (Retinopie) immer weiter zugunsten von hochspezialisierten Zentren hin verlassen. V1 und V2 sind annähernd noch vollständig retinop aufgebaut. In den höheren Kortexarealen hingegen gibt es hochspezifische Zellen, die zum Beispiel nur auf das Vorhandensein von Händen, Tieren oder Gesichtern reagieren, unabhängig davon, wo diese Objekte sich im Sehfeld befinden.
Wird ein zuvor unbekanntes Objekt wahrgenommen, wird der feiner aufgelöste Bereich der Augen (die Fovea centralis) darauf gerichtet und das Objekt optisch abgetastet. Hat bereits eine Kategorisierung stattgefunden, wird das Objekt sofort erkannt (Perzeption) und auch die Wahrnehmung selber wird diesem Objekt angepasst. Dies geht sogar soweit, dass Kantenwahrnehmung von Scheinkanten, also nicht vorhandenen Kanten, auch im visuellen Kortexareal V1 stattfindet, tatsächlich vorhandene Kanten hingegen dort dann nicht mehr ausgewertet werden. Handelt es sich hingegen um ein völlig unbekanntes Objekt, wird es genauer betrachtet und dann einer passenden Kategorie hinzugefügt (Apperzeption). Ist eine solche Einteilung erfolgt, findet nur noch eine Perzeption statt. Bei einer Fehlbeurteilung dauert es daher recht lange, bis eine korrigierende Wahrnehmung möglich ist.
Die tatsächliche, physikalische Auflösung eines Auges ist auf wenige Bogensekunden begrenzt. Durch die neuronale Fusion beider Bilder beim stereoskopischen Sehen wird tatsächlich eine wesentlich höhere Auflösung wahrgenommen (etwa um Faktor 10 besser), als sie der Bauplan des Auges erahnen lässt. Diese Überauflösung (engl. hyperacuity) hat mehrere Ursachen, wie die neuronal feinere Auflösung im V1 und die langsamen Augenbewegungen (drift). Die schnelle Auswertung der Signale der Photorezeptoren und die Retinopie bis in den visuellen Kortex führt auch häufig zu optische Täuschungen. Die Farbwahrnehmung entspricht der Standard-Farbwahrnehmung aller Wirbeltiere (Farbkreis), obwohl diese eigentlich für vier verschiedene Photorezeptoren ausgelegt zu sein scheint.
Die Verknüpfung der visuellen Wahrnehmung mit anderen Wahrnehmungen geschieht über jeweils eigene Hirnareale. So werden akkustische Wahrnehmungen vermutlich über den Colliculus inferior mit der visuellen Wahrnehmung verknüpft. Zur Ermittlung der Lotrechten wird neben der Information des Innenohres auch der wahrgenommene Horizont verwendet. Daher "ziehen" Abgründe den Betrachter an, da eine aus beiden Informationen gemittelte Lotrechte wahrgenommen wird.
Quelle: WIKIPÄDIA
zurück ...